









Введение
В различной литературе можно встретить утверждение, что пирамиды оказывают специфическое воздействие на различного рода процессы – физические, химические, биологические и т.д. Этот так называемый «эффект пирамид» ныне не находит объяснения в рамках известных законов, поскольку (по утверждению его исследователей) воздействие оказывает сама форма пирамиды как таковой. Причем, по утверждению тех же исследователей, «эффект пирамид» максимален в случае соблюдения нижеследующих условий:
- угол наклона граней пирамиды равен углу наклона граней Великой пирамиды на плато Гиза в Египте;
- пирамида сориентирована по сторонам света;
- объект воздействия располагается в геометрическом центре пирамиды (эффект чуть меньше в центре ее основания).
Поскольку целый ряд публикаций об «эффекте пирамид» вызывает обоснованные сомнения в достоверности их утверждений, в ходе экспедиции в Египет, организованной под эгидой Фонда развития науки «III тысячелетие» в декабре 2009 года, проведена серия экспериментов. Цель экспериментов – натурная проверка реальности наличия самого «эффекта пирамид», в чем бы он не заключался.
В частности, проведенные эксперименты включали в себя и такие, которые ставили целью зафиксировать воздействие пирамид на биологические процессы.
В рамках биологического эксперимента семена гороха (Pinum sativum) сорта Немчиновский были разложены по группам в лабораторных условиях. Одна группа оставалась в г.Пущино Московской области, остальные с 29.11.09 по 14.12.09 находились в Египте.
Условия хранения и воздействия:
1 группа: хранилась в Пущино при температуре 10-140С
2 группа: была перевезена в Египет, но оставалась в отеле (данная группа семян служила контролем)
3 группа: семена находились в Великой пирамиде на плато Гиза в предкамере (геометрический центр пирамиды) в течение 7 суток
4 группа: семена находились в пирамиде-спутнице Великой пирамиды G1a в ее камере (примерно чуть ниже центра основания пирамиды) в течение 7 суток
5 группа: семена находились в дальней камере Красной пирамиды в Дашуре (примерно центр основания пирамиды) в течение 2 суток.
1. Измерение всхожести семян
18.12.09 в 12.30 семена были разложены в специальные растильни (см. рис. 1) и проращивались в лаборатории при комнатной температуре 20-230С. Динамику проклевывания семян наблюдали примерно каждые 2 часа до момента пока не проклюнуться все семена. Через сутки, т.е. в 11.30 19.12.09 все семена были набухшие, но не было ни одного проклюнувшего семечки. Первый замер был сделан в 14.20, т.е. через 26 часов. Семечко считали «проклюнувшим» когда появлялся белый кончик корешка. Результаты исследования представлены на рис. 2.

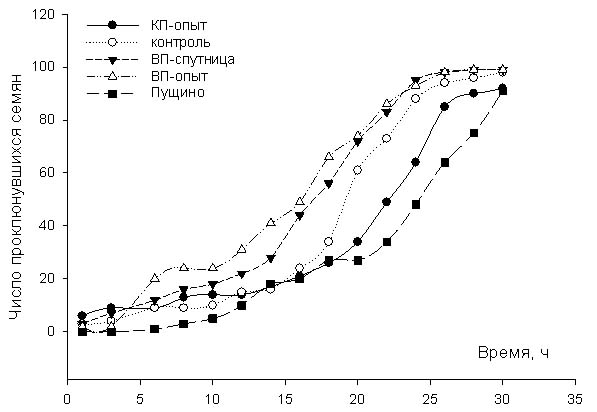
На рис. 2 и 3 видно, что через 56 часов все семена проклюнулись. Однако, динамика проклевывания отличается у разных групп. Не вызывает удивление, что динамика проклевывания групп семян, находившихся в Египте отличается от динамики проклевывания семян, находившихся в Пущино. Известно, что на всхожесть семян влияют условия, в которых они находились до момента набухания. Т.е. повышенная температура и, в большей степени, повышенная влажность ускоряют выход семян из состояния покоя. Данные, представленные на рис. 2 подтверждают этот факт. Однако, вызывает удивление, что всхожесть семян, находившиеся в Красной пирамиде близка к всхожести семян, находившихся в Пущино.

Примечания. КП- Красная пирамида, ВП-Великая пирамида, СВП- спутница Великой пирамиды. Общим контролем считаем семена, находившиеся в гостинице. В графе «время» первое значение обозначает время с момента первого замера, значение в скобках – время с момента замачивания семян в растильне.
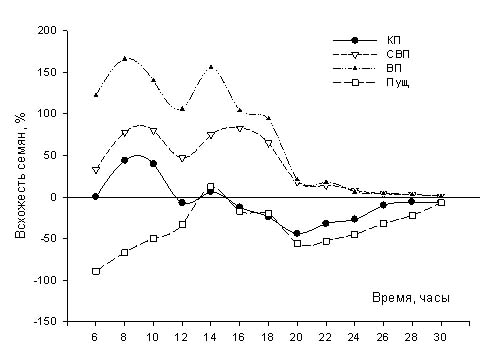
Для сравнения стоит привести кинетику прорастания семян, облученных в оптимально стимулирующих дозах радиации, 10-20 сГр.
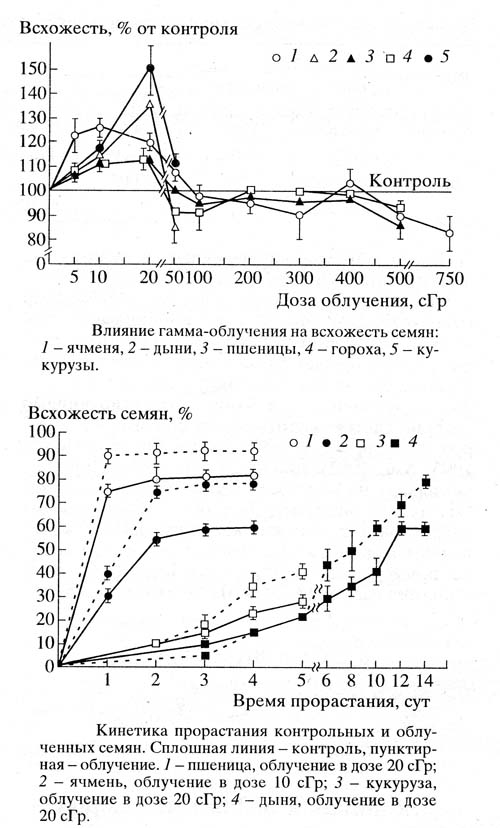
Дозы радиации 5-20 сГр считаются малыми дозами радиации (для сравнения повреждающими дозами считаются от 2 до десятков Гр). В цитируемой работе показано, что при таких дозах происходит стимуляция всхожести семян, в том числе и семян гороха. При дозах радиации 50-400 сГр эффект отсутствует и подавление всхожести семян наблюдается при дозах свыше 500 сГр. Исследование кинетики прорастания семян при дозах 10-20 сГр позволяет авторам сделать вывод, что облучение увеличивает вероятность выхода семян из состояния покоя и не влияет на скорость процессов, приводящих к их проклевыванию (набухание, структурные и метаболические изменения в тканях зародышей).
В нашем случае мы наблюдаем отсутствие эффекта на кинетику проклевывания в Большой пирамиде и некоторое подавление для семян, находившихся в Красной пирамиде. Уместно ли в данном случае говорить об изменении уровня радиации – сказать трудно.
Для справки
Прорастание семян делится на фазы набухания семян, проклевывания, гетеротрофного роста, перехода к автотрофному питанию. Попадание семян во влажную среду выводит их из состояния покоя. Поглощение воды является пусковым фактором прорастания. Это поглощение осуществляется благодаря повышению проницаемости семенных покровов для воды и за счет гидратации биополимеров в клетке. В результате развивается онкотическое давление (давление набухания) и семенные покровы разрываются. Набухание практически не зависит от температуры, содержания О2, освещения. Проклевывание начинается, когда семена достигают критической влажности (40-65% в пересчете на сырую массу), и происходит путем роста растяжением самого зародышевого корня, в результате чего кончик корешка выталкивается из семени. После этого начинается рост за счет деления клеток. Выход корня обеспечивает закрепление прорастающего семени в почве (в субстрате) и улучшает поглощение воды. Вслед за корнем начинается рост побега.
2. Измерение длины проростков гороха
После роста корня начинается рост наземных органов – стеблей, а затем формирование листьев. Проростки находились в одинаковых условиях: при одинаковой температуре, влажности и искусственном освещении. (Следует отметить, что декабрь не самое подходящее время для проращивания гороха, поскольку в это время семена должны находиться в физиологическом покое).
Измеряли проростки обычной линейкой с точностью до ± 0.1 см два раза в сутки. Данные представлены на следующих графиках:
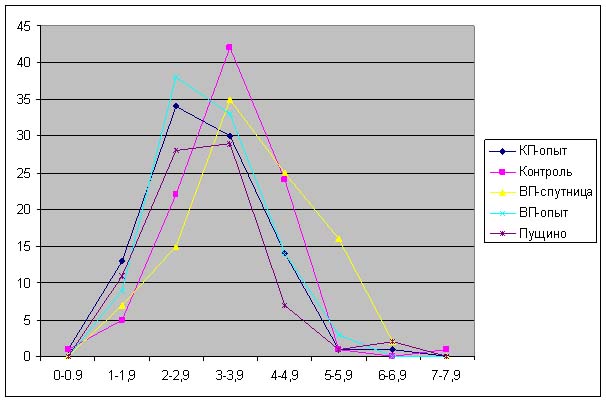
Большая часть длин проростков, полученных из семян, которые находились в Красной и Великой пирамидах лежит в диапазоне от 1.5 до 4.0 см, в то время как длина проростков, выращенных из контрольных семян – от 2 до 4.9 см (рис. 5). Распределение длин проростков, выращенных из семян, находившихся в пирамиде-спутнице близко к контрольному значению.
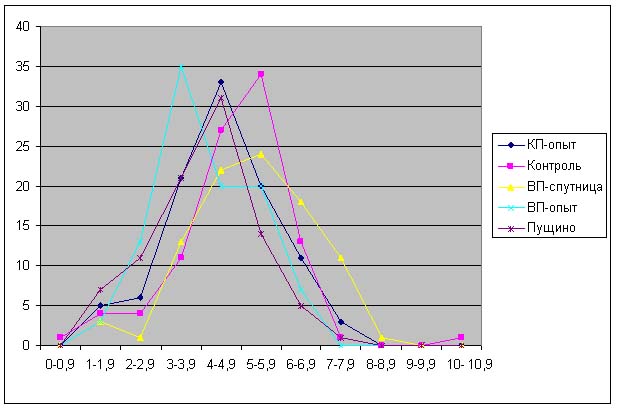
На рис. 6 можно наблюдать, что увеличивается «расхождение» между распределением длин контрольных проростков и проростков, выращенных из семян, находившихся в Великой пирамиде. При этом распределение длин проростков из семян, находившихся в Пущино совпадает с распределением длин проростков, выращенных из семян, находившихся в Красной пирамиде.

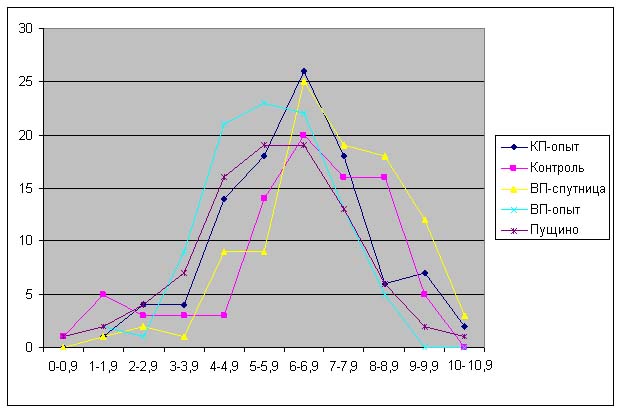
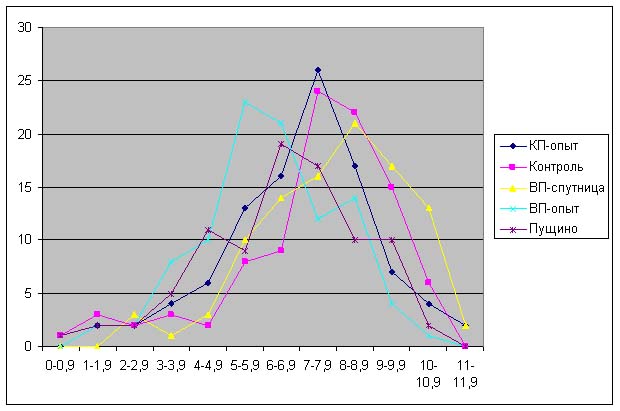
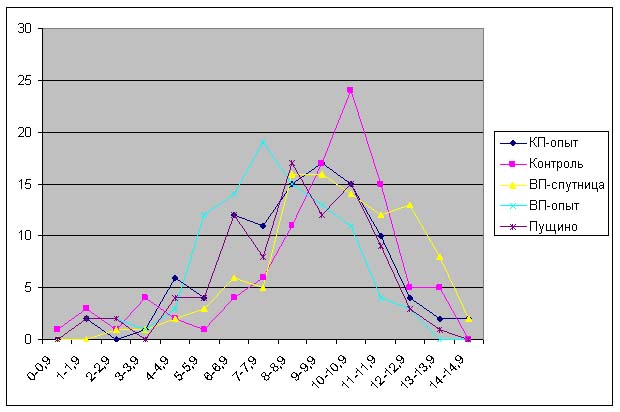
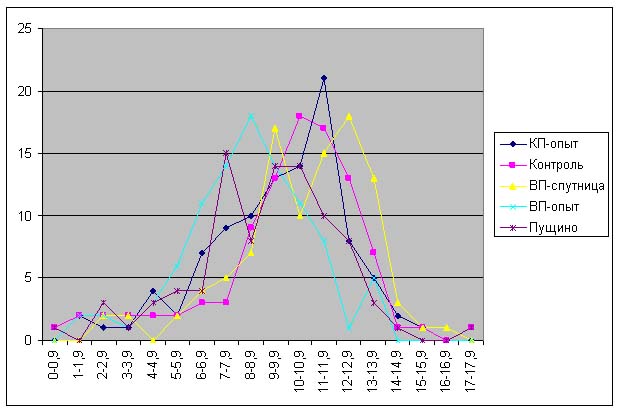
Анализ данных, приведенных на рис. 5-11 показывает, что проростки гороха, выращенных из семян, являвшихся контролями, растут несколько быстрее, чем проростки гороха, пророщенные из семян, находившихся в самих пирамидах. Кроме того, следует отметить, что в некоторых случаях скорость роста проростков из семян, находившихся в пирамидах близки к скорости роста проростков из семян, находившихся в Пущино. Напомним, что семена находились в Пущино при температуре 10-140С, а также другой влажности и другой длине светового дня. Исходя из вышесказанного, однозначный вывод о влиянии пирамид на прорастание семян был бы преждевременным. В данном случае, можно отметить лишь тенденцию к замедлению прорастания проростков из семян, находившихся в пирамидах.
3. Исследование продуктов перекисного окисления липидов в хлоропластах листьев проростков гороха
Многочисленные исследования последних десятилетий показали, что независимо от природы воздействия, ответ растения на него развивается по некоторой общей схеме, что позволяет говорить о существовании неспецифической стрессовой реакции на воздействия извне. Таким универсальным звеном в реакции растительного организма на действие самых разнообразных факторов может быть некоторое стереотипное изменение внутренней среды клетки, на роль которого многие исследователи выдвигают окислительный стресс. Большое внимание в последнее время уделяется механизмам ответа живых систем на слабые внешние воздействия. Предполагается, что действие факторов, по силе не выходящих за зону толерантности организмов, формируется не вследствие прямого повреждающего эффекта стрессора, а в результате разбалансирования процессов повреждения и восстановления клеточных структур. В связи с этим исследование слабых воздействий может способствовать выявлению механизмов тонкой регулировки внутренних процессов, происходящих при внешних возмущениях. В клетке постоянно протекают окислительно-восстановительные процессы, способные приводить к появлению активных форм кислорода (АФК). Наиболее эффективно этот процесс идет на сопрягающих мембранах митохондрий и хлоропластов. Хлоропласты являются теми структурами растительной клетки, в которых образование АФК происходит с наибольшей эффективностью. В силу своей высокой активности, АФК способны вызывать повреждение важнейших биополимеров: нуклеиновых кислот, белков, липидов и углеводов. Поэтому неотъемлемой чертой всех аэробных организмов является неферментативное окисление биологических молекул по радикальному механизму с участием кислорода. АФК генерируют органические гидропероксиды ROOH, подобные по структуре пероксиду водорода и также химически активные. При последующем метаболизме они переходят в спирты, альдегиды, эпоксиды и т.д. Этот процесс называется перекисным окислением, а совокупность этих реакций – окислительными модификациями молекул.Однако наиболее активно свободнорадикальное окисление идет при атаке инициирующим радикалом диаллильного атома углерода, в связи с этим в реакцию наиболее активно вступают полиненасыщенные жирные кислоты липидов мембран. В растениях окислению подвергаются преимущественно линолевая, линоленовая и олеиновая кислоты. Первичными продуктами перекисного окисления липидов являются пентадиенильные радикалы, образующие диеновые конъюгаты и гидроперекиси жирных кислот. Дальнейшее их окисление и распад приводит к образованию широкого спектра конечных продуктов: альдегидных и спиртовых производных с укороченной цепью, в частности 4-гидрокси-2-ноненаль, низкомолекулярные продукты (этан, пентан), малоновый диальдегид, эпоксиды. Выход окислительных процессов из-под контроля может быть губителен для клетки, однако в то же время существует представление об АФК, как о нормальных и даже необходимых метаболитах аэробной клетки. В общем случае ПОЛ является универсальным модификатором свойств биологических мембран, важным физиологическим регулятором их структуры и функций, фактором, устанавливающим и поддерживающим стационарное функционирование ферментов, каналообразователей, рецепторов.
Методы исследования
Приготовление суспензии хлоропластов
Суспензию готовили методом дифференциального центрифугирования, основанном на разности коэффициентов седиментации разных органелл клетки (Arnon et al., 1956).
Навеску листьев гороха 1 г растирали с 7 мл среды выделения (10,5 мл 1 М NaCl + 6 мл 0,1 М трис-HCl буфер + 14,5 мл воды) в фарфоровой чашке во льду. Разливали в центрифужные пробирки и центрифугировали 6 мин при 4 тыс. об./мин при охлаждении (4˚С). Супернатант сливали, осадок (хлоропласты) ресуспензировали с 1,5-2 мл 0,035 М NaCl.
Определение содержания диеновых конъюгатов
Степень диеновой конъюгации ненасыщенных высших жирных кислот определяли по методике И.Д. Стальной (1977).
Для определения диеновых конъюгатов 0,5 мл суспензии растворяли в 4,5 мл смеси гептан: изопропиловый спирт в соотношении 1:1 и центрифугировали 10 мин при 4 тыс. об/мин. К надосадочной жидкости добавляли 0,1 объема воды для разделения фаз. К 0,5 мл верхней гептановой фазы добавляли 2,5 мл этанола и спектрофотометрировали против контроля (гептан : спирт в соотношении 1 : 5). В ходе ПОЛ на стадии образования свободных радикалов в молекулах НЖК возникает система сопряженных двойных связей, что сопровождается появлением нового максимума в спектре поглощения при 232 нм.
Расчет ДК производили по формуле:
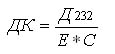
где Д232 – оптическая плотность, Е – коэффициент молярной экстинкции, 2,1.10-6 моль-1. см-1, С – концентрация липидов, мг / мл.
Диеновые конъюгаты выражали в мкМоль ДК / мг липидов.
Определение липидов
Содержание липидов проводили спектрофотометрически по реакции с фосфорно-ванилиновой смесью.
Обычно продукты перекисного окисления измеряют в хлоропластах проростков (в случае, когда речь идет о растениях), подверженных какому-либо воздействию, например, радиации. По этому поводу существует большое количество работ. Однако, в проростках, выращенных из семян, подвергшихся какому-либо воздействию, продукты перекисного окисления не меряют. Особых оснований полагать, что уровень продуктов перекисного окисления каким либо образом поменяется в таких проростках, нет. Тем не менее окислительно-восстановительные процессы протекают на мембранных структурах, в том числе на мембранах хлоропластов, протекают постоянно за счет образования активных форм кислорода (АФК). Результатом увеличения продукции АФК, как правило, является смещение прооксидантно-антиоксидантного равновесия в сторону интенсификации окислительных процессов, об активности протекания которых можно судить по накоплению продуктов ПОЛ. В некоторых работах ПОЛ рассматривается как маркер неспецифического ответа организма на воздействие извне. ПОЛ – цепной процесс свободнорадикального окисления – приводит к образованию пентадиенильных радикалов, которые образуют диеновые и триеновые конъюгаты и гидроперекиси жирных кислот. Конечными продуктами ПОЛ являются альдегидные и спиртовые производные гидроперекисей, низкомолекулярные продукты (этан, пентан), эпоксиды, малоновый диальдегид (МДА).
В нашей работе мы измеряли диеновые конъюгаты (первичные продукты ПОЛ) и основания Шиффа, которые образуются на более поздних стадиях. Разумеется, для того чтобы определить окислительно- восстановительный статус клетки, нужно промерить большее количество продуктов ПОЛ, а также ферменты антиоксидантной системы, которые поддерживают окислительно – восстановительный баланс. Однако, задачей данного исследования было посмотреть будет ли проявляться различие нахождение семян в разных условиях на продукты ПОЛ, образующихся в проростках из таких семян.
Результаты
Красная пирамида – разница по отношению к контролю:
ДК: -18% ± 7%
ОШ: -21% ± 2%
Великая Пирамида – разница по отношению к контролю:
ДК: 14%±4%
ОШ: 16%±3%
Спутница к Великой пирамиде – разница по отношению к контролю:
ДК: 3%±12%
ОШ: 4%±3%
Контроль в Пущино – разница по отношению к контролю:
ДК: -12% ± 8%
ОШ: -9%±1.5%
Великая пирамида – разница по отношению к Пирамиде – спутнице:
ДК: +15% ± 16%
ОШ: +11%± 3%
Красная пирамида по отношению к контролю в Пущино:
ДК: +7% ± 13%
ОШ: -9% ± 1.5%
Великая пирамида по отношению к контролю в Пущино:
ДК: +24% ± 11%
ОШ: +32%±3%
Спутница к Великой пирамиде по отношению к контролю в Пущино:
ДК: +7% ± 13%
ОШ: +20%+3%
4. Электронно-микроскопическое исследование прилистников проростков гороха
Данные электронно-микроскопического исследования прилистников проростков гороха представлены на рис. 12-16.
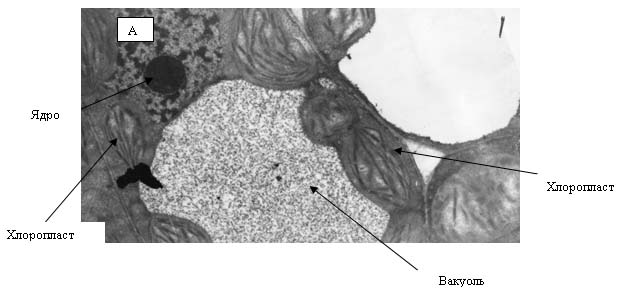
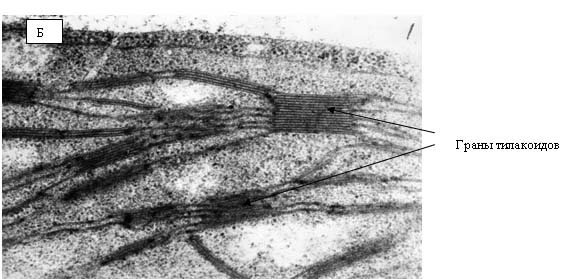
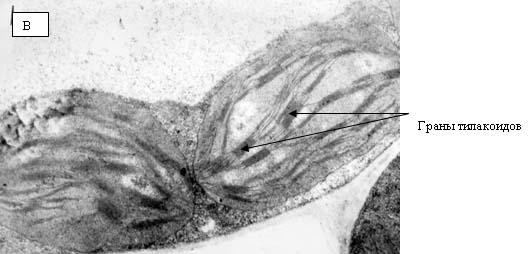
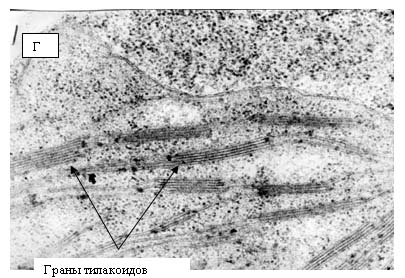
Рис. 12. Электронные фотографии клетки прилистников проростков гороха, полученных из семян, находившихся в Красной пирамиде.
На рис. 12 и 13 представлены электронные фотографии, выполненные при исследовании ультраструктуры прилистников проростков гороха, выращенных из семян, находившихся в Красной пирамиде (рис. 12) и контрольной группы (рис. 13). На рисунках видно, что никакой разницы в ультраструктуре между упомянутыми группами растений не наблюдается. На фотографиях четко видны граны тилакоидов, которые расположены ровными «столбиками» в строме хлоропласта. Следует отметить, что именно таким образом эти органеллы выглядят «в норме».
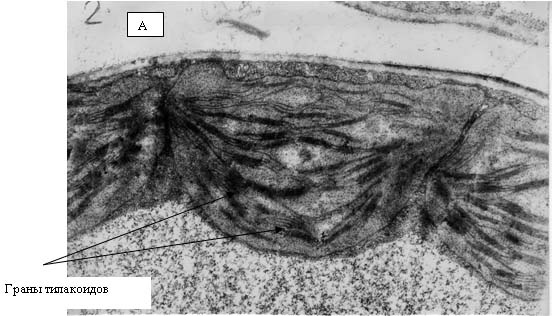
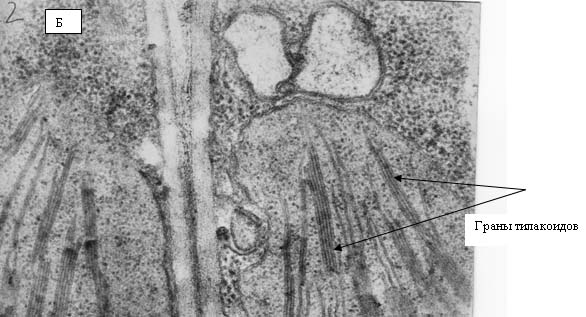
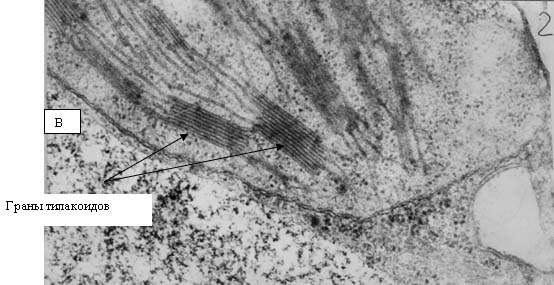
Рис. 13. Электронные фотографии прилистников проростков гороха, полученных из семян, являвшихся общим «контролем»

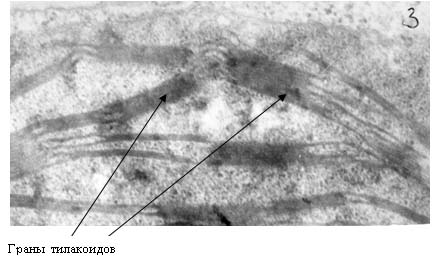
Рис. 14. Электронные фотографии прилистников проростков гороха, полученных из семян, находившихся в пирамиде – спутнице к Великой пирамиде.



Рис. 15. Электронные фотографии прилистников проростков гороха, полученных из семян, находившихся в Великой пирамиде.
Ультраструктура клеток прилистников проростков гороха, полученных из семян, находившихся в Великой пирамиде (рис. 15) и пирамиде – спутнице (рис. 14) отличаются от проростков, выращенных из семян, находившихся в Красной пирамиде и контрольной группе. На рис. 14 видно, что клетки прилистников в пирамиде – спутнице характеризует наличие пустот в цитоплазме, набухшие, но пустые митохондрии, тилакоиды хлоропластов плотно сжатые. Следует отметить, что митохонодрии – это органеллы клеток, на мембранах которых происходит ряд окислительно – восстановительных процессов, в результате чего образуется молекулярный кислород. Внутренняя мембрана митохонодрий большое количество инвагинаций, что обычно хорошо видно на электронных фотографиях (но никак не пустот). Что касается ультраструктуры клеток прилистников, полученных из семян, находившихся в Великой пирамиде, то на рис. 15 также можно наблюдать гетерогенность цитоплазмы со множествами пустот. Что касается тилакоидов хлоропластов, то можно наблюдать как набухшие тилакоиды (рис. 15-б), так и сильно сжатые тилакоиды (рис. 15-в). Стоит отметить, что ультраструктура клеток прилистников, выращенных из семян, находившихся в Великой пирамиде близка к ультраструктуре клеток прилистников проростков гороха, выращенных из семян, находившихся в Пущино – такая же гетерогенная структура цитоплазмы и наличие как сильно набухших тилакоидов (рис. 16 –б), так и сильно сжатых (рис. 16-в).
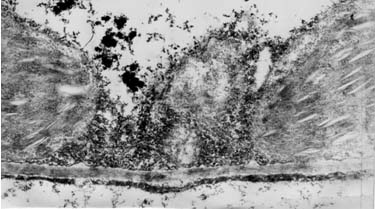
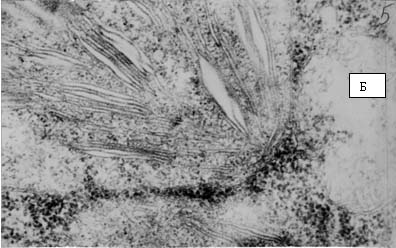

Рис. 16. Электронные фотографии прилистников проростков гороха, полученных из семян, находившихся в Пущино
Заключение
Все представленные выше результаты не позволяют сделать однозначного заключения о том, что нахождение семян в Красной или Большой пирамидах могут изменять морфологические, биохимические или функциональные характеристики растений, выращенных из этих семян. Для проверки достоверности каждого полученного результата, нужно провести еще как минимум 4-5 экспериментов. Однако, суммируя все перечисленные данные можно предположить о некоем «намеке» на эффект биологического действия пирамид. Однозначный вывод трудно сделать еще и потому, не известно находились ли семена контрольной и опытной групп в одних одинаковых условиях по температуре и влажности. Кроме того, учитывая перевозку семян в страну с другими климатическими условиями, необходимо моделировать соответствующие условия (температура, влажность) для контрольной группы в России (например, в Пущино). Еще один немаловажный фактор – время года. Возможно, что результаты, полученные, в марте – сентябре, резко отличались бы от приведенных результатов экспериментов, выполненных в декабре (время физиологического покоя). Что касается самих физических факторов, которые могли бы оказывать влияние на семена, то можно выделить следующие:
1. Изменение постоянного магнитного поля Земли (полное или частичное экранирование).
2. Изменение (полное или частичное экранирование переменной составляющей) естественного магнитного поля.
3. Радиактивный фон.
4. Аэроионы (радиолиз воздуха).
Первые три пункта не вызывают сомнения, а в качестве пояснения к по п.4 ниже приводится отрывок из статьи, опубликованной в Сборнике трудов IV Конгресса «Сверхслабые поля в биологии и медицине».
И еще одно замечание, хорошо известно, что совместное действие, например, двух различных факторов может вызывать аддитивные эффекты, а может проявляться в отсутствии биологического эффекта, как например, химические вещества плюс магнитное поле или магнитное поле плюс действие малых доз радиации. Поэтому, даже в данном случае отсутствие видимых биологических эффектов не позволяет делать заключение, что пирамиды не оказывают влияние на биологические системы – в нашем случае на семена и проростки, выращенные из таких семян.
Приложение
Введение из статьи В.В. Леднев, Н.А. Белова, Л.К. Сребницкая «Влияние парафинового экрана на свойства дистиллированной воды и раствора карбоната натрия».
Основными источниками ПРФ на поверхности Земли (на уровне моря) являются радиоизотопы (U, Ra, Rn, Th) и продукты их распада, входящие в состав почвы, воды, воздуха (внешние источники) и живых организмов (внутренние источники). К внешним источникам облучения относятся и вторичные галактические космические лучи (ГКЛ), также постоянно воздействующие на живые организмы. Согласно различным оценкам, относительный вклад вторичных ГКЛ в суммарную годовую дозу облучения составляет 20-30%.
К настоящему времени показано, что ПРФ необходим для нормального развития животных, растений и микроорганизмов. Снижение как внешнего, так и внутреннего естественного облучения, сопровождается ингибированием скорости деления простейших и сине-зеленых водорослей, снижением скорости прорастания растений из семян, снижением скорости развития насекомых и снижением скорости роста (набора веса) молодых мышей и крыс. В большинстве из перечисленных экспериментов снижение ПРФ достигалось с помощью использования свинцовых экранов толщиной 5-10 см или, реже, при размещении тест-систем в помещениях, расположенных на большой глубине под земле. Биоэффекты, полученные в перечисленных выше опытах интерпретировались как результат понижения внешнего и внутреннего ПРФ.
Очевидно, что размещение тест-систем в свинцовых экранах, шахтах и под толстым слоем воды должно сопровождаться не только подавлением радиоактивного фона изотопов (внешних источников ПРФ), но и снижением воздействия вторичных ГКЛ. Тем не менее, ни в одной из соответствующих работ возможный вклад ГКЛ в функционирование биологических систем не рассматривается. Можно предположить, что из числа различных вторичных частиц ГКЛ наиболее вероятными возможностями биотропного воздействия могут обладать нейтрон. Это предположение находит косвенное подтверждение в результатах некоторых лабораторных исследований.
Анализ статистических данных показал, что достоверное увеличение регистрируемого числа инфарктов и инсультов наблюдается во время снижения интесивности вторичных нейтронов на поверхности Земли, (так называемые Форбуш – понижение интенсивности ГКЛ]. Проведенные нами ежедневные в течение 3-х месяцев измерения сорбции водопроводной воды сухими семенами гороха, также выявили корреляцию между Форбуш – понижениями и усилением сорбции воды семенами гороха [неопубликованные данные].
Особый интерес представляют экспериментальные данные, свидетельствующие о том, что экранирование сосуда с дистиллированной водой парафиновыми и свинцовыми экранами сопровождается значительным (на 20-30 %) увеличением порога ультразвуковой кавитации воды. Согласно авторам этих исследований, наблюдаемый эффект обусловлен, главным образом, изменениями физических характеристик вторичных нейтронов ГКЛ после их прохождения через экран окружающий сосуд с водой. На возможность корреляции свойств воды с некоторыми геофизическими индексами указывают также некоторые другие публикации.
Как известно, энергия вторичных нейтронов ГКЛ, достигающих поверхности Земли, существенно понижается при их прохождении через материалы, содержащие высокую концентрацию водорода – парафин, дерево, полиэтилен, вода. Вместе с тем, характер воздействия нейтронов на различные материалы, в том числе, и на биосистемы существенно зависит от энергии нейтронов.. Естественно предположить, что взаимодействие вторичных нейтронов ГКЛ с биосистемами может измениться после их прохождения через водород – содержащий экран. Для проверки этой гипотезы мы предприняли изучение влияния парафинового экрана на свойства некоторых физико-химических и биологических систем.


