










Чтобы понять каким конкретным образом эволюционирует живой мир, произведем дифференциацию всей живой биологической массы по бросающимся в глаза способа контактирования с внешней средой (с реальным объектом). Таких способа два – вегетационный и сенсорный. К вегетационному способу контактирования принадлежит растительный мир, к сенсорному (чувственному) – животный. Сенсорный контакт животного мира с внешней средой может быть или чисто инстинктивным или инстинктивно разумным. Палеонтологические данные убеждают нас в том, что эволюционирует и растительный мир и животный. Не вдаваясь в подробности этих данных и не соотнося их с существующей научной дифференциацией геологического времени, но обобщая эти подробности феноменологически, вернемся к нашей работе “Мы – мутанты” [3, с. 147-153] и рассмотрим следующую схему:
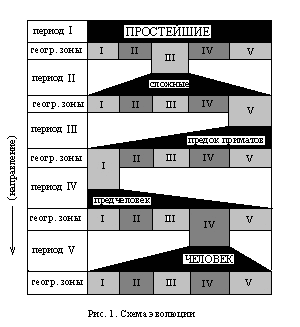
“Представим себе нашу планету упрощенно. Допустим, что есть на ней всего лишь ять географических зон, И пусть на нашей планете появились простейшие… Их появление знаменует собой начало I периода.
Отсутствие конкурентной среды (золотой век!) позволило им беспрепятственно освоить все разнообразие имеющихся на планете географических зон (в нашем случае – условные пять). Но разнообразие этих зон детерминировало генетический отбор в соответствующем направлении, что в итоге привело к тому, что простейшие каждой зоны стали приобретать свои отличительные признаки, свойственные только им. Конечным результатом такого процесса было образование первых видов простейших (в нашем случае: условные пять зон – условные пять видов). Один из этих видов в результате такой эволюции приобрел более широкий спектр приспособительных функций, чем другие – превратился из простейшего в нечто более сложное и допустим, что это произошло в III географической зоне. Отметим также и ту очевидность, что филогенетическое преобразование простейших в некое видовое многообразие привело постепенно к тому, что межвидовое размножение стало невозможным, а если даже и да, то этому мешали новоприобретенные физиологические особенности, заставлявший каждый индивид, в силу выживания, гнездиться рядом с таким же, как и он – т. е. колонизировать среду обитания. Далее логика событий рисует нам следующую картину. Сложные, обособившиеся в III зоне, как более сильные и более выживаемые, начали мигрировать в остальные зоны, вытесняя и уничтожая своих собратьев. Вытеснение и уничтожение не приводило к полному истреблению – оно означало лишь то, что хотя остальные виды и продолжали параллельное существование, эволюционно-генетическое преобразование их значительно замедлялось.
После распространения сложных по всем пяти географическим зонам (которых в действительности было значительно больше) процесс эволюционно-генетического отбора повторился. Оговоримся, что и этот, II период филогенеза мы значительно упростили, как по горизонтали (количество географических зон), так и по вертикали, ибо он содержит в себе не менее одиннадцати периодов (проторезойский, кембрийский, силурский, девонский, карбонский, пермский, триасский, юрский, меловой, третичный и четвертичный)…
Представим себе, что благодаря случайному стечению обстоятельств сильнейшими оказались сложные, живущие в V зоне. Назовем тот вид животных, в которых в этой зоне превратились сложные, предком приматов. И, действительно – пройдя в течение многих миллионов лет через ряд эволюционно-генетических преобразований, они могли в конце третичного периода в какой-либо из географических зон (например, в одном из районов Восточной Африки) превратиться именно в такое животное. И оно, будучи предком приматов, превосходя другие виды в находчивости, ловкости, хитрости и сплоченности, начало экспансивную миграцию из V зоны в остальные и, расселившись, начало эволюционировать в соответствии с вариациями внешней среды.
В одной из зон (допустим в I) предок приматов превратился в вид, который мы условно назвали предчеловеком (III период филогенеза). И, наконец, еще одна метаморфоза привела к тому, что предчеловек превратился в человека (IV зона, IV период).
В настоящее время (V период) практически нет такого места на земле, где бы не обитал человек. Период этот еще не закончился и он резко отличается от характера эволюционно-генетических процессов прошлых периодов филогенеза. Прежде всего тем, что несмотря на то, что фенотипические формы эволюции успели привести человека к расовым, национальным и прочим видам межзонального различия, генотипические ее формы ведут к увеличению мощи и силы разума, который в противовес агрессивности и антагонистичности не дал и не дает человеку дифференцироваться на несовместимые виды, ибо в любом случае способствует… …наследственно-генетическому взаимопроникновению”.
Здесь мы описали феноменологически течение эволюции во времени в соответствии с одной из многих существующих научных гипотез. Надо сказать, что схематически описанный нами процесс вполне укладывается в рамки дарвинизма и сегодня никого удивить не может. И конечно же не ради того, чтобы у кого-нибудь от удивления поднялись брови, мы предпринимаем очередное наше путешествие в незнаемое. И мы, конечно же, не намериваемся опровергать Дарвина. Эволюция есть и этот факт не подлежит никакому сомнению. Однако наряду с неопровержимостью самого этого факта, пугает не столько многообразие гипотез, пытающихся объяснить конкретные пути эволюционного процесса, сколько возможность их легкого опровержения. Ну, например, одна из таких гипотез, утверждает что в результате быстрого погружения дна океана, многие животные оказались вдали от береговой кромки, что изменило само по себе условия их существования настолько, что они мгновенно вымерли и на смену им пришли новые виды, сумевшие приспособиться и выжить. Когда начинаешь интересоваться сколько понадобилось геологического времени для такого быстрого опускания и мгновенного вымирания, то оказывается, что не менее 50000 лет. Теоретически такого временного промежутка вполне достаточно, чтобы мышь в результате эволюционных преобразований могла превратиться (в смысле размеров) в слона, а такое животное, как например, кит – в мышь. Но если это так, то возможно ли объяснять массовую гибель животных (палеонтологически доказанную) и возникновение новых видов изменением условий существования? Строго мыслящие исследователи, принимая во внимание продолжительность катастрофы, выдвигают более подходящую гипотезу, как например, уважаемый нами Д. Соболев. Он утверждает следующее: “Возможность обращения развития обходит значительную часть тех препятствий, какие возникают при отыскании родоначальных форм различных филетических ветвей ныне живущих и ископаемых организмов не среди фантастических и нежизненных схем, а между действительно существовавшими живыми и специализованными формами. Однако, одна эта возможность сама по себе еще не устраняет всех затруднений. Если бы обратное развитие происходило медленно и постепенно, в таком случае мы легко находили бы в ископаемом состоянии следы таковой деградации в виде постепенных переходов между теми группами организмов, которые относятся друг к дружке, как предки к потомкам. В действительности мы этого не наблюдаем. Факты палеонтологии не укладываются в рамки кунтактивной эволюции – прямой или обратной” – [5, с. 133]. Под кунтактивностью здесь понимается постепенность процесса. Конкретизируя свою мысль до наглядного представления того как может осуществляться такое возвращение специализированных организмов к примитивным формам, Соболев предполагает, что в филогении явление обратного развития проявляется в том, “что в филетическом ряду каждый последующий член заканчивает свое онтогенетическое развитие – в отношении всех или только некоторых признаков – на более ранней стадии, чем предшествующий член: в последовательном ряду поколений потомки “неотенически” не дорастают до тех стадий, на которых стояли предки” – [5, с. 104, 105]. Под неотенией в данном случае разумеется возможность более раннего развития и полового созревания.
С таким предположением ученого, несмотря на то, что он приводит много палеонтологических примеров, можно и не соглашаться, исходя из доводов пригодных для абсолютно противоположной точки зрения, но и плюс к этому достаточных для защиты иных гипотез, имеющих такое же право на существование, как и представленная Соболевым. Если отбросить его гипотезу, как неподтвержденную более убедительными фактами, чем теми, которые подчеркиваются в его работе (например, “что птеридоспермы (Cycadofilices) появились задолго до возникновения настоящих папоротников, так что высший тип исторически предшествует низшему” – [5, с. 110], то что остается? Наличие существенных результатов наблюдений над современными популяциями, экстраполяция которых на более длительные периоды времени убеждает в том, что эволюция на протяжении миллионов лет скорее всего происходила путем постепенных прогрессивных изменений. Однако данные палеонтологической летописи, особенно по морским беспозвоночным, свидетельствуют о параллельных возможностях совершенно иных процессов. Элдредж и Гулд (Eldredge, Gould, 1972) отметили, что среди ископаемых форм, лишь очень немного надежно документированных случаев существенных изменений вида с течением времени. Наиболее обычные из них сохраняют свои характерные морфологические особенности на протяжении миллионов лет. Когда же изменения все-таки наступают, они, насколько можно судить по находкам, требуют относительно короткого периода по сравнению с общей продолжительностью существования вида. И это практически то единственное, что нам достоверно известно. Не потому ли, например, располагая практически одной и той же информацией, одни ученые приходили к выводу, что внутри вида Homo erectus объем черепной коробки существенным образом не менялся (Rightmire, 1981), тогда как другие (Cronin et al., 1981) настаивали на постепенных изменениях в ряду от Australopithecus до Homo sapiens?
Исходя из отмеченного нами спорного опыта предшественников, сразу же оговоримся, что, внося свой скромный и, весьма вероятно, далеко не академический вклад в гипотетические построения апостолов научной мысли, мы не собираемся опровергать ни одной из существующих гипотез. Очень возможно, что все они (не станем утомлять читателя их перечислением) имеют вполне законное право на существование – хотя бы по той явной причине, что эволюция в способах восхождения от низшего к высшему не имеет ограничений в выборе путей развития. Подобная недетерминированность характерна, прежде всего, для позвоночных. И связано это очевидно с геометрической особенностью первого деления оплодотворенного яйца. Если у специализированных форм различных групп беспозвоночных, включая кольцевых червей, членистоногих, и головоногих моллюсков первые деления яйца происходят так, что клетки (бластомеры) следующего по счету ряда вклиниваются между клетками предыдущего (это называется спиральным дроблением) и, таким образом, судьба каждой клетки предопределена с самого начала, т. е. если один из бластомеров удалить, соответствующая ему часть организма попросту не разовьется; то у хордовых и иглокожих, родоначальников всех существовавших и существующих ныне позвоночных, “первые клеточные деления протекают по типу радиального дробления, т. е. бластомеры располагаются точно друг над другом. Если одну из клеток на ранних стадиях онтогенеза удалить, соседние могут компенсировать ее недостачу и разовьется нормальный зародыш (недетерминированное деление)” – [4, т. I, с. 30].
Нам кажется, что выход хордовых и иглокожих в эволюционном процессе на недетерминированное деление клеток стал возможным только потому, что биохимическое образование правых и левых молекул в живых организмах характеризуется несимметричностью (впервые замечено Пастером), которая сдвигает любую живую систему в сторону неустойчивости и посему делает ее максимально способной к новым формообразованиям в соответствии с изменениями внешней среды.
Уделяя чрезвычайное внимание этой несимметричности, В.И. Вернадский отмечает в связи с этим неравенство правых и левых спиралей в форме организмов, выявляемое “в редком проявлении “левшей”, причем для некоторых организмов левши являются преобладающими и могут быть принимаемы как видовой признак” – [2, кн. 1, с. 58]. Здесь следует отметить замеченную нами идентичную несимметричность в мире насекомых и в мире самых высокоразвитых животных, проявляемую в расположении органов воспроизводства звука. У кузнечиков, например, правое надкрылье находится под левым. Указуя на эту природную закономерность, Фабр Ж. А. В своей знаменитой книге “Нравы насекомых” пишет – “смычок левого надкрылья есть двигатель; терка правого надкрылья – точка колебания; натянутая кожица зеркальца – резонатор, вибрирующий при посредстве своей приводимой в сотрясение рамки” – [6, т.II, с. 388].
Источник порождения звуковых сигналов несимметричен и у китов. “Направленность посылаемого сигнала контролируется специальным образованием, расположенным на голове около ноздри. У современных одонтоцетов это одно отверстие ( у остальных китов их два), и его модификация приводит к заметной асимметрии окружающих костей. Такая асимметрия возникла независимо шесть, а то и десять раз при формировании различных семейств одонтоцетов. Можно сгруппировать зубатых китов в пять надсемейств, соответствующих крупным филогенетическим радиациям. Каждое надсемейство характеризуется особым типом асимметрии” – [4, т. III, с. 94].
После наших искренних уверений, что мы не собираемся опровергать ни одной из имеющихся гипотез в области эволюционного изменения видов зачем, собственно говоря, нами затеян весь этот разговор? Конечно же не для того, чтобы удивить читателя эрудицией, ибо, очевидно, что какой бы обширной она не казалась, она черпает информацию из готовых источников знаний, а наша задача внести в эту копилку нечто новое. Исследуя ту проблему, в которую мы, со всей очевидностью, уже углубились и проанализировав сложившуюся ситуацию, мы можем с уверенностью сказать, что все существующие гипотезы выражают только одну сторону какого либо палеонтологически фиксируемого явления и посему одна гипотеза не походит на другую и ей противоречит. Исходя из этого положения, бросающегося в глаза, назрело, вероятно, время для такой теоретической основы, которая бы тяготела не к индивидуальной оригинальности, но стремилась бы удовлетворить все прежние гипотезы – т. е. не являлась бы по отношению к ним противоречивой. Для того, чтобы приблизиться к поставленной цели необходимо определить недостаток всех предшествующих теорий и недостаток этот должен быть в равной мере присущ без исключения им всем.
Нам кажется, что все существующие гипотезы, относящиеся к путям и возможностям эволюционных преобразований, страдают односторонним подходом к проблеме, заключающемся в том, что рассматривается сам факт морфологического изменения и предполагаемое соответствующее влияние внешней среды, и, исходя из этого плоскостного подхода фантазируется та или иная возможность, когда сущность явления заключается в том, чтобы ответить на коренной вопрос – что происходит с изменяющимся организмом внутри, каков порядок эволюционного преобразования – изменение внешнего воздействия (условий среды обитания) и в результате последующее приспособительное изменение организма, или, как мы предполагаем, существует третий фактор – промежуточный и самый важный – процесс реагирования организма, который необходимо рассматривать в самой его материальной сущности?
Расшифруем то, что мы имеем ввиду более наглядно. Если мы говорим слово “деревья”, то мы всего лишь обозначаем и обобщаем некоторую определенную сумму реалий внешнего мира. Но внутренняя сущность самого дерева остается при этом нам абсолютно неизвестной. Наложим эти наши рассуждения на такое выражение: эволюция организмов есть результат реагирования их на изменение внешней среды. Все вроде бы верно сказано, но если первая часть этой логической формулировки подтверждается всевозможными палеонтологическими находками и внешняя среда может изменяться в наглядной степени, то, что такое “реагирование” в самой своей сущности? Быстрое отдергивание руки при прикосновении к раскаленному предмету не есть само реагирование, но всего лишь внешний результат каких-то глубинных процессов, происходящих в организме. И для того, чтобы верно ответить на вопросы, относящиеся к эволюционным преобразованиям, нам необходимо докопаться до этих глубинных процессов – увидеть “реагирование” в глубинно-предметной его сущности. И опять же, если мы уже заглянули “внутрь” этого процесса, то сразу же оговоримся, что нас здесь не интересуют моменты и способы сенсорного восприятия, в том числе и способы передачи поступившего из реальности сигнала по нервным волокнам и даже не само вместилище этих сигналов (нервные узлы – ганглии, и мозг в том числе). Нас интересует конкретная точка воздействия и даже не столько она, а именно то, что происходит внутри живого организма, когда сигнал достигает конечного своего пункта и какие побудительные представления и мотивы порождаются при этом в самом организме – будь это человек, насекомое или любое другое животное.
Для того, чтобы упростить вхождение в поставленную проблему, начнем с насекомых, но не потому, что строение у них более простое, чем, скажем, у позвоночных, но потому именно, что согласно существующим научным воззрениям, поведением насекомых руководит только и только инстинкт. Подчеркнем здесь слово “поведением”, ибо нас интересует прежде всего поведение, поскольку оно выражает содержание инстинктов. выработанных в течение миллионов лет. И поскольку мы ни коим образом не являемся специалистами в области энтомологии, руководящим источникам для нас в этой научной области будут наблюдения и записки Ж. А. Фабра, известного своей знаменитой книгой “Инстинкт и нравы насекомых”.
Ученый описывает свои наблюдения в подробностях, представляющих для нас тематический интерес – “…церцерис выбирает места с плотной, утрамбованной, твердой почвой, …места эти должны быть сухи и лежать на солнечном припеке. В этом выборе обнаруживается такой разум или, если хотите, инстинкт, что я склонен считать его результатом опыта” – [6, т. I, с. 14]. Церцерис – название одного из видов ос. Или, например, описанное Фабром поведение церцерис бугорчатой: “Если какая-нибудь пластинка песчаника выдается в виде карниза, если в почве встречается естественное углубление величиной с кулак, то насекомое пользуется этим навесом или этим углублением для устройства под ним своей галереи, прибавляя таким образом естественные сени к своей постройке” – [6, т. I, с. 18]. И еще – в том же ключе: “Оса становится лицом к лицу с своей жертвой, схватывает ее хобот своими могучими челюстями, и в то время, как долгоносик выгибается на своих ножках, оса передними лапками усиленно давит его в спину, …чтобы раскрыть сочленение брюшка… …и… …в два или три приема впускает свой ядовитый стилет в место прикрепления передней части груди, между первой и второй парами ножек” – [6, т. I, с. 26] . Здесь Фабр описывает каким точнейшим и отработанным способом оса парализует нервные центры, ответственные за систему движений и долгоносик тут же становится неподвижным. Примечательно, что оса охотится только на пластинчатоусых жуков (долгоносиков и златок), нервная централизация которых организована таким образом, что этих жуков можно парализовать одним уколом. Удивленный результатами собственных наблюдений, ученный восклицает: “По совершенству инстинкта и нравов перепончатокрылые не только занимают первое место среди насекомых, но превосходят значительно и многих высших животных” – [6, т. I, с. 562]. Внимательное прочтение труда Фабра наталкивает нас на вывод, что насекомые действую только и только в пределах ритуала свойственного в особенностях своих каждому отдельному виду. “Для инстинкта нет ничего трудного, – говорит Фабр [6, т. I, с. 83], – до тех пор, пока действие не выходит из обычного круга деятельности, отведенного животному; для инстинкта также нет ничего легкого, если действие должно отклониться от обыкновенного пути. Насекомое, которое удивляет и поражает нас своей проницательностью, минуту спустя, перед фактом самым простым, но чуждым его обыкновенной практике, удивляет нас своей тупостью. И далее следует описание конкретного наблюдения за поведением одного из видов ос – “…ограбленный сфекс входит к себе и осматривает пустую комнату, а минуту спустя ведет себя так, как будто бы он не заметил исчезновения объемистой добычи… Он замечает все это. Но в таком случае, зачем эта другая глупость, заставляющая его закупоривать очень добросовестно пустую ячейку, которую он и позже не собирается снабжать провизией? Труд запирания здесь совершенно бесполезен и в высокой степени бессмыслен. Значит различные инстинктивные поступки насекомых фатально связаны между собой и два действия их находятся в такой зависимости одно от другого, что совершение первого влечет за собой совершение второго даже тогда, когда вследствие случайных обстоятельств второе сделалось не только ненужным, но иногда даже противоречащим интересам насекомого” – [6, т. I, с. 87-88]. И все же знаменитый энтомолог не отказывает насекомым в некоторой поведенческой свободе выбора, но все это в рамках раз и навсегда выработанного ритуала, в пределах которого обозначена сфера деятельности насекомого. В подтверждение этого Фабр рассказывает: “Халикодома только что окончила первый слой крышечки на ячейке и улетела за другим комочком земли для дальнейшего укрепления крышечки. В ее отсутствие я протыкаю иглой крышечку и делаю в ней широкую брешь, занимающую половину отверстия. Насекомое возвращается и прекрасно починяет пролом. Будучи занято устройством крышечки, оно при починке ее продолжает ту же работу” – [6, т. I, с. 405]. И затем, рассказывая об искусно варьируемых способах вмешательства в текущую работы насекомого, Фабер резюмирует результаты экспериментирования: “В то время насекомое только начинало строить и мое вмешательство коснулось работы, которой пчела занималась в тот момент; сделанная мной дырочка явилась в глазах пчелы как бы погрешностью в ее постройке, которая легко может случиться в откладываемых слоях, еще не успевших высохнуть. Поправляя эту погрешность, каменщица не оставляла начатой работы. Но раз начался принос провизии, начальная ячейка считается оконченной и, чтобы не случилось, пчела не коснется ее больше. Теперь заделать дыру значило бы изменить порядок работы, а этого насекомое не может сделать. Теперь очередь меда, а не цемента. На этот счет правило непоколебимо” – [6, т. I, с. 408]. Можно было бы и закончить приводить наглядные примеры ритуального поведения насекомых, но больно уж интересно описывает свои наблюдения Ж. А. Фарм. Вот, например, еще одно описание – речь идет о ритуально-инстинктивном поведении жука-навозника, именуемого испанским копром, а именно о самке – о присущем ей материнском инстинкте: “Мать… проявляет полное самоотречение: она отказывается от радостей свободы и простора, от ночных полетов и от рытья в свежем навозе… Спрятавшись под землей, возле семьи, она не выходит больше из своей детской, сторожит ее, счищает паразитную растительность, замазывает щели и устраняет всякого появляющегося хищника и паразита… В сентябре она выходит на поверхность со своей семьей, которая, не нуждаясь в ней больше, скоро отделяется и живет отныне по-своему. Птица не проявляет более преданного материнства” [6, т. II, с. 73].
Отметим по ходу предпринятого нами путешествия, что ритуальность поведения присуща не только одному индивидуальному насекомому, а всему виду. И несомненно, что каждый представитель вида хорошо различает своих и чужих. “Самое замечательное, – говорит Фарм о пчеле осмии, – глубокое почтение к соседнему, еще не взломанному, кокону. Как бы осмия не спешила выйти, она не тронет его челюстями: это священная вещь. Она разломает перегородку, с остервенением станет грызть стену, даже если там останется одна древесина, она все вокруг себя обратит в прах; но тронуть стесняющий ее кокон она никогда себе не позволит” – [6, т. I, с. 366]. Заметив такую преданность себе подобным Фабер проделывает следующий эксперимент: “В стеклянную трубочку или в сложенные желобки ежевичного стебля я кладу, попеременно, коконы осмия и коконы соления (солений бродячий – Solenius vagus – прим. наше). Результат получается поразительный. Осмии, как и следует, вылупляются раньше и тотчас выходят из гнезда, причем они превращают коконы соления и их обитателей в клочки, где я ничего не могу разобрать, кроме разве валяющихся там и сям отдельных голов. Итак, осмия не пощадила живых коконов другого вида… …животное совершенно равнодушно ко всему, что не ее племя” – [6, т. I, с. 368]. В связи с таким, присущем насекомому голосу крови, очевидно, что оно имеет тягу к сосуществованию с себе подобными. И хотя на первый взгляд у многих из них не существует никакой общины, они любят гнездиться вместе. Наблюдая поведение осы церцерис бугорчатой, Ж. А. Фабр говорит о насекомых этого виды: “Хотя у них не существует никакой общины, но эти насекомые любят гнездиться вместе в небольшом числе: я наблюдал их гнезда всегда группами, по меньшей мере до десяти” – [6, т. I, с. 19]. Или, например, такой момент, отмеченный ученым: “Щетинистая аммофила до сих пор встречалась мне в равнинах, всегда поодиночке, и живет она так же в одиночку… …а потому я был очень удивлен, найдя на вершине Ванту это перепончатокрылое собравшимся в таком большом количестве под камнем” – [6, т. I, с. 110]. Изучая жизнь пчелы халикодомы амбарной, ученый-энтомолог приходит к выводу, что это насекомое индивидуалист – работает только на себя и не заботится о других. Но тем не менее предпочитает жить в большом обществе (устраивает целые колонии). Содружество в этом пчелином многолюдии проявляется только тогда, когда все ячейки готовы и яйца снесены – “тогда пчелы, не различая своих ячеек от чужих, начинают делать общий покров для всей колонии, состоящей из толстого слоя цемента, который заполняет промежутки между ячейками и прикрывает всех их вместе” – [6, т. I, с.319]. Наблюдение и изучение жизни насекомых привели Фабера к факту существования у них памяти и адекватного зрительного восприятия реальности. В связи с этим ученый повествует об осе помпиле. Он сообщает, что добычей этому насекомому служит паук. Перед тем как рыть норку, оса оставляет паука неподалеку от роемой норки на листке облюбованного ею куста, и затем возвращается к своей добыче. Ученый экспериментирует – перекладывает паука на открытое место. Оса возвращается к ранее облюбованному кусту и не находит паука. Ищет и заметив паука, переносит его на лист другого куста. Перелетает к норке, которую продолжает рыть и через некоторое время проверяет находится ли добыче там, где она ее оставила. Она летит прямиком к этому последнему кусту – к тому листику, на котором был оставлен паук. Фабр поражен памятью этого карапуза. Знаменитый энтомолог утверждает, что осе “достаточно один раз, второпях, увидеть какое-нибудь место, ничем не отличающееся от других, чтобы хорошо вспомнить его, несмотря на свои ревностные занятия земляной работой” – [6, т. I, с. 260]. Но ученый на этом не успокаивается и делает пальцем небольшое углубление в земле, куда кладет паука и прикрывает тоненьким листиком. “Тогда оса мучается в поисках пропавшей дичи, проходит несколько раз через листик, не подозревая, что под ним лежит паук, и продолжает свои поиски дальше. Значит ею руководит не обоняние, а именно зрение” – [6, т. I, с. 260]. Вот к какому интересному выводу приходит ученый. И здесь возникает конкретный вопрос – если насекомые обладают памятью, то в тот момент, когда они что-то вспоминают, образное это воспоминание, или какое либо другое? Предположим, что воспоминание это носит образный характер. И поскольку насекомые руководствуются в своем поведении только инстинктом, то из нашего предположения вытекает, что инстинкт заложен в них в виде передающихся по наследству образных матриц, определенный набор которых является неотъемлемой частью каждого вида насекомых и образные эти матрицы в отличие от присущего человеку образных матриц коллективного бессознательного, можно назвать видовым бессознательным. И если все то, что мы здесь гипотетически предполагаем, действительно выработано у насекомых, то поведение их зависит от набора этих матриц и очередности доминирования во времени матричных образов, которые предопределяют инстинктивный порядок и особенности поведения насекомых того или иного вида в процессе жизненного цикла.
И конечно, не стоило бы в таком объеме приводить моменты из наблюдения Ж. А. Фабра, если бы то, к чему мы пришли в результате его наблюдений и экспериментов, не имело бы прямое отношение к эволюционным преобразованиям. Мы утверждаем следующее – появление новой матрица (архетипа, если хотите) побуждает организм к новым преобразованиям. При неизменном наборе этих архетипов никакое воздействие внешней среды не может привести ни к генотипическому изменению, ни к фенотипическому. При возникновении нового архетипа (нового видового бессознательного) в некоем сообществе любого вида животных эволюционные преобразования могут иметь место даже в том случае, если условия среды обитания уже не те в сравнении с теми, которые привели к возникновению этого нового архетипа.
Но здесь возникает вопрос – каким образом проявляется рождение нового архетипа? Ответ один – в изменении ритуального поведения. При этом необходимо учитывать, что изменение такое может наступить только в том случае, когда оно присуще не индивидууму, а некоей группе, которая в количественной сумме своей (критическая масса) достаточна для того, чтобы отречься от прежнего ритуала и начать новую жизнь.
Отметим, что резкое изменение видового бессознательного (ритуального поведения) явление настолько редкое, что описание такого явления мы нигде, даже у самых знаменитых естествоиспытателей, не встречали – исключение в этом ряду принадлежит только Ж. А. Фабру. Описав это событие, ученый не придал ему особого значения, ибо ему и в голову не пришло привязать его к эволюционному преобразованию. Но вот интереснейшее по своей значимости свидетельство самого ученого: “В тот момент, как сфекс совершает свой визит в норку, я беру сверчка, оставленного у входа, и кладу его несколькими дюймами дальше. Появляется сфекс, издает свой обыкновенный крик, с удивлением глядит туда и сюда и, видя, что дичь слишком далеко, выходит из норки, чтобы схватить ее и опять положить в прежнем положении. Сделав это, он опять спускается в норку, но один. Прежний маневр повторяется с моей стороны и то же разочарование со стороны сферкса. Но дичь опять принесена им ко входу в нору и насекомое опять сходит одно; все одно и то же до тех пор, пока мое терпение не истощилось. Раз за разом я повторял мой опыт сорок раз над одним и тем же сфексом; его упорство победило мое, а тактика его никогда не изменялась.
В течение некоторого времени это непоколебимое упорство, найденное мною у всех сфексов одной колонии, не переставало беспокоить мой ум. Я говорил себе: значит, насекомое повинуется фатальной склонности, которую ни в чем не могут изменить обстоятельства; его действия неизменно однообразны и ему чужда способность приобрести из собственных действий хотя бы малейшую опытность.
Новые опыты изменили этот слишком абсолютный взгляд. Спустя год я посетил то же самое место. Новое поколение унаследовало для норок новое место, выбранное предшествовавшим поколением, а также неизменно унаследовало его приемы: опыт с отодвиганием сверчка дает те же результаты. Заблуждение мое все увеличивалось, когда счастливый случай натолкнул меня на другую, отдаленную, колонию сфексов. Здесь я опять принимаюсь за те же опыты. После двух или трех раз, когда результат был прежний, сфекс садится на спину найденного сверчка, схватывает его челюстями за усики и немедленно втаскивает в норку. Кто остался в дураках? Экспериментатор, которого перехитрила умная оса. Соседи его, у других норок, где раньше, где позже, так же точно пронюхивают мои хитрости и без остановок вносят дичь в свое жилье,. вместо того чтобы упрямо оставлять его на пороге. Что это значит? Поселение, которое я изучаю теперь, …более искусное, чем поселение прошлого года” – [6, т. I, с. 45-46].
Если насекомым свойственно искать общество себе подобных, то тем более это свойственно многим видам животных, для которых групповой образ жизни является наследственной чертой, выработанной в борьбе за существование. Инстинкт хищных рыб собирает их вместе, как бы группирует – только таким образом они могут успешно окружать добычу, отбивать одну из рыб от стаи, оттеснять ее от убежищ, прижимать к кромке воды, дезориентировать ее и сигнализировать друг другу о нахождении добычи.
Поведение животных в стае строго регламентировано и носит оно зачастую ритуально-гипнотический характер. “Понаблюдаем за стайкой рыбок в аквариуме. Одна из них плывет впереди, остальные следуют за ней растянувшись цепочкой. Но вот передняя рыбка достигает стенки, стайка, подобно солдатам в строю, повернулась “кругом” и бывший лидер стал замыкающим. Стайку ведет другая рыбка, и следующие за ней повторяют все ее повороты, обходят камни с той же стороны, почесывают бока о те же выступы” – [1, с. 36]. О ритуально-гипнотическом характере поведения животных говорит и такая любопытная информация: Если напугать стадо, олени сначала соберутся в более плотную массу, а потом направятся за первым же выскочившим из стада оленем. Он в период такого гона будет играть роль вожака. Описанное здесь поведение стада имеет автоматический, инстинктивный характер, поскольку в подобных случаях оно постоянно повторяется – является выработанной формой ритуального поведения. Результаты экспериментальных методов показали наличие 18 – 30% таких потенциальных вожаков в любом достаточно крупном стаде. “Если средний размер стада принять в 2000 голов, то примерно 600 животных в нем способны к самостоятельному, активному поведению в неблагоприятной обстановке” – [1, с. 39]. Или такие, например, достоверные сведения: “В одной отаре из 600 животных лишь четыре овцы решились выйти из отары и, глубоко увязая в снегу, увлекли за собой остальных. Те же самые четыре вожака, по словам чабана, водящего эту отару 7 лет, способны повести овец вплавь через бурную горную реку. Самостоятельность этих овец проявилась без всякого научения, сразу же, когда им было не более года от роду” – [1, с. 43].
Влияние в стаде вожака беспредельно. Подчинение вожаку всех членов стада иначе как гипнотическим не назовешь. В домашнем овцеводстве известны случаи, когда за случайно сорвавшимся вожаком устремлялись все остальные члены стада. Исходя из таких случаев, описанная в Евангелии гибель свиней выглядит вполне правдоподобно:”Паслось же там при горе большое стадо свиней. И просили Его все бесы, говоря: пошли нас в свиней, чтобы нам войти в них. Иисус тотчас позволил им. И нечистые духи, вошедши, вошли в свиней; и устремилось стадо с крутизны в море, а их было около двух тысяч; и потонули в море” – (Мар. 5, 11-13). Что здесь, собственно говоря, описано, как не явление массовой гибели животных гипнотического порядка. Хорошо известны и неоднократно зафиксированы случаи стадного самоубийства китов – они выбрасываются на берег и если из милосердия вернуть их в эти гибельные моменты в родную стихию (такие эксперименты были проделаны), то они возвращаются и повторяют начатое, пока не погибают. До сих пор такое поведение китов остается загадкой – причину приписывают нарушению психического состояния животных, вызванное влиянием магнитных аномалий. Но представим себе такую картину. Вожак китового стада внезапно “съехал с катушек”. Вместо обычных ритуально-поведенческих архетипов у него, по каким-либо причинам (испуг или, скажем, голод – отсутствие в рационе любимого планктона) стал доминировать архетип времен его предки кондилартров – абсолютно сухопутных животных. Тоска по тем библейским для него временам обуяла его настолько, что кит решил вернуться на сушу. И поскольку он вожак, то и остальные киты, гипнотически завороженные его сумасшествием, последовали за ним. Не так ли в принципе происходило массовое вымирание животных в палеонтологические эпохи. Нет, мы не шутим – мы говорим об этом вполне серьезно.
Реконструкция скелета верхнеюрской птицы археоптерикса (Arhaeopteryx) говорит о типе очень близком к свойственному мелким целурозавровым динозаврам типа Compsognatus с массой около 3,5 кг. Это было, если судить по скелету, мелкое быстро бегающее животное. Весьма вероятно, что основной пищей ему служили насекомые (в том числе и летающие). В мозгу этого динозавра, постоянно занятого охотой зрительно отпечатался архетип возможности полета – это животное хорошо знало и помнило, что живность за которой он охотится способна передвигаться по воздуху. Кроме того известно, что этот тип мелкого динозавра относился к чешуйчатым. И вполне вероятно, что во время быстрого бипедального бега чешуя могла топорщится по воле животного на определенных участках тела и в особенности на передних лапах, которыми ящер сбивал добычу. Во время погони за насекомым с помощью чешуи менялось встречное сопротивление воздуха и резкие повороты выполнялись успешнее, чем у гладкокожих животных. А если надо было прыгать с какого либо уступа вниз за улетающим насекомым, то в такие моменты чешуя становилась парусом, увеличивая дальность прыжка и плавность спуска. Но здесь надо заметить, что чешуя эта никогда не стала бы превращаться в перья, если бы этот динозавр “не свихнулся” на возможности полета – т. е. вначале появляется доминирующий архетип, а потом соответствующее эволюционное изменение. Но и это еще не все. Если эти животные жили группой. И групповое их общение было ритуально обозначено и среди этих динозавров, появился индивид, у которого наметилось превращение чешуи в перья и передних конечностей в крылья, то вряд ли, благодаря такому достоинству, он мог стать вожаком группы. Это могло произойти только в том случае, если бы не он один мог совершать более дальние прыжки и быть ловким в беге, но и другие – способные вслед за ним совершить точно такой же прыжок с обрыва, какой удавался ему, а иначе последовавшие его примеру погибли бы, или, в лучшем случае, были бы искалечены. Размышление приводит нас к тому, что в стаде (в группе) должно появиться критическое количество особей, чтобы одна особь из них могла возглавить всю группу и предложить новую ритуальную форму поведения. Возникновению особей с новыми генетическими признаками всегда привязано к появлению критического количества таких особей и к генетически наследственной передаче нового архетипа из поколения в поколение. Отметим, что накопление таких новых признаков процесс довольно медленный и морфологические изменения при этом могут быть настолько незначительными, что в ископаемых остатках они зафиксированы быть не могут. Но вот в группе появилось достаточно особей с новыми генетическими признаками для того, чтобы навязать большинству новую ритуальную форму поведения. Что в этот момент происходит? Есть два варианта. Первый – те особи, которые не могут по своим генетическим возможностям вписаться в новую форму ритуала, мгновенно вымирают, что и зафиксировано палеонтологической летописью. Те, которые могут его выполнять, освободившись от примитивно сдерживающего фактора – эволюционно сметенных с дороги сородичей, в течение буквально нескольких поколений реализовывают архетип-доминант – окрыляются буквально и действительно становятся способными к полету. Второй вариант отличается от первого тем, что особи не вписывающиеся в новый ритуал, отделяются от “бунтовщиков”, выбирают вожака из своих и продолжают филетическую линию развития – “эволюционируют по Дарвину”.
Первый вариант преобразований является революционным. Попытаемся его изобразить графически:
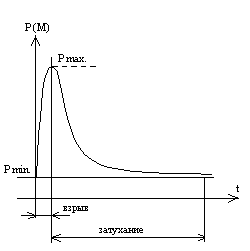
По оси абсцисс на рис. 2 отложено время, по оси ординат – величина морфологического изменения P. Здесь Р max – максимальная величина морфологического изменения (начало революционного морфологического изменения сведено к нулю). P min – минимальная величина морфологического изменения (присуща виду постоянно). Посему, практически, революционное изменение начинается не с нуля, а с P min.
С чисто математической точки зрения скорость изменения в нашем случае является отношениемΔP/Δt – т. е. отношением приращения бесконечно малого морфологического изменения в течение временного промежутка, соответствующего этому приращению, к величине этого временного промежутка В соответствии с нашей гипотезой кривая скорости эволюционных преобразований экспоненциально распределена в отрезке времени от революционного взрыва (толчка) до момента превращения этого взрыва в кунтактивно текущий эволюционный процесс. Внутренний смысл явления заключается в том, что после взрыва происходят быстрые морфологические преобразования (внешне выражаемые, например, в превращении чешуйчатого покрытия в оперение и передних конечностей в крылья); но после того, как преобразование завершилось, процесс быстро затухает, ибо архетип, вызвавший взрывное преобразование был реализован во всех направлениях и морфологически и в ритуальных формах. В дальнейшем новый вид ортодоксально придерживается новых ритуальных форм – точно так же, как вид, от которого он произошел длил когда-то или продолжает свое эволюционное деление в рамках старого ритуала. Этим самым мы желаем сказать, что видовое бессознательное тяготеет к постоянству и устойчивости более, чем к новым архетипическим приобретениям.
В связи с вышеприведенными рассуждениями и соображениями, правомерность применения единицы для измерения скорости эволюции, именуемой дарвин, вызывает у нас большое сомнение, впрочем, как и всякая иная условно придуманная величина.
Еще один интересный вопрос, касающийся кривой скорости эволюционных преобразований. Нам думается, что законы описанные нами и действующие в животном мире, распространяются и на человеческие сообщества в одинаковой мере со времени появления первого человека до наших дней. Как эти законы наглядно проявляются? – спросит дотошный читатель. Ответ: в постоянных революциях и бунтах, сопровождающих известную нам длительность исторического процесса. Что есть эти бунты и революции, как не попытка сломить установленную систему иерархий, а значит и ритуальных отношений? Неудачи всех этих попыток заключается лишь в том, что в обществе не накоплено достаточное количество особей с доминированием бунтующего архетипа и, как только это произойдет – наступит одна единственная революция, которая сметет начисто иерархическую установку наживы, издевательства над личностью и подавления творческих устремлений. Осуществление такое станет возможным потому, что у человека, в отличие от животных, разум соотносится с инстинктом со знаком равенства и даже занимает главенствующее положение. А это означает, что наступит время, когда не эволюционные процессы будут управлять развитием человеческих сообществ, а наоборот человек будет абсолютным их хозяином. К этому устремлению его будет понуждать необходимость завоеваний космических пространств и потребность в связи с этим мгновенного приспособления к новым – непредвиденным условиям существования.
Возвращаясь от наших фантазий к узкому направлению избранной нами темы, отметим, что зафиксированные палеонтологической летописью факты массового вымирания животных в том случае, если эти вымирания связаны с планетарными или космическими катастрофами, вносят в эволюционный процесс ощутимые помехи и этим самым со всей несомненностью влияют на дальнейшую скорость и направление этого явления, но ни в коем случае не выражают закономерностей процесса – закономерностей, которые, как мы это выяснили, лежат внутри самой эволюции. Попробуем развернуть наше рассуждение конкретнее.
Период границы между мелом и третичным периодом называют сокращенно границей К-Т. Ее называют также маастрихт-датской границей по последнему меловому и первому третичному ярусам соответственно. В районах этой границы, охватывая по толщине слоя как плюс так и минус, отмечено крупномасштабное вымирание животных и растений. И поскольку в горизонте К-Т-границы почти во всех исследованных местах обнаруживаются четкие пики иридия, то, исходя из этого факта, возникло несколько гипотез причин глобального вымирания. Суть одной из этих гипотез заключается в том, что на землю в те далекие времена упал астероид диаметром в 10 км. и в результате иридиевое облако заволокло верхние слои атмосферы и послужило препятствием для освещения земной поверхности солнечными лучами. На земле наступили долгая ночь и резкое похолодание. Выжили только мелкие млекопитающие, типа грызунов, которые будто бы и послужили источником новых эволюционных преобразований. А выжили они в таких условиях, благодаря интенсивному метаболизму, позволившему им в катастрофически экстремальных условиях “сохранять активность в течение более длительных периодов времени и поддерживать высокую, постоянную температуру тела, независимую от внешней среды” – [4, т. II, с.197].
Мы не исключаем возможности того, что катастрофа на границе К-Т действительно имело место, как и долгая ночь (в сетчатке примитивных современных млекопитающих палочки количественно преобладают над колбочками!), как и похолодание, как и то даже, что предками человека являются именно эти грызуны, пережившие глобальную катастрофу. Мы утверждаем лишь то, что все предполагаемые катастрофические явления оказывают на процесс эволюции лишь побочное действие, ибо эволюция эта происходит и без катастроф во всей полноте по законам непреложно ей присущим. Главным источником эволюционных преобразований является возникновение нового архетипа в видовом бессознательном и генетическое его распространение и накопление до появления в стаде критического числа видов, носителей этого нового архетипа – этой “сумасшедшинки” с поведенческой точки зрения носителей рутинного ритуала. И посему наша мысль сводится к тому, что динозавр может научиться летать только в том случае, если инстинктивное желание осуществить такую возможность будет архетипически закреплено в нем задолго до полета.
Список литературы:
1. Баскин Л. М., Законы стада, издательство “Знание”, Москва 1971. – с 48.
2. Вернадский В. И., Размышления натуралиста, пространство и время в неживой природе, в 2 кн., изд. “НАУКА”, Москва 1975, кн. I – 1975 – с. 176, кн. II – 1977. – с. 192.
3. Кобринский А., Мы – мутанты, Беер-Яков 1996. – с. 256.
4. Кэрролл. Р., Палеонтология и эволюция позвоночных, в 3-х т., пер. с англ. – М: “Мир”, I т. – 1992 – с. 280, II т. – 1993 – с. 283, III т. – 1993 – с. 312.
5. Соболев Д., Начала исторической биогенетики, Гос. изд. Украины 1924. – с. 205.
6. Фабр Ж. А., Нравы насекомых, в 2-х т, пер. с фран., – М: “ТЕРРА” – “TERRA”, 1993, I т. – с. 608, II т. – с. 612.



Добавить комментарий